Феномен и ноумен времени. Том 1 (1), 2004. С. 16-19.
© Г. А. Голицын, 1992
Принцип максимума информации в эволюции материиГ. А. Голицын |
||
Москва, МИРЭА Эволюционный подход почти одновременно сложился в физике и в биологии, однако опирался на разные принципы и предполагал разную направленность эволюционного процесса. Согласно физическому принципу максимума энтропии, любая замкнутая система (а возможно, и мир в целом) стремится к максимальной неупорядоченности, к разрушению любых структур. В биологии, напротив, эволюционный подход призван был объяснить, каким образом в ходе развития возникают все более сложные и упорядоченные системы и структуры. Таким образом, встает проблема: как может возникать упорядоченность в мире, где энтропия в целом возрастает? Если не считать виталистических концепций, исповедующих ту или иную разновидность "жизненной силы", особого фактора, действующего только в живых системах и объясняющего их специфику, то ответа на эти вопросы нет до сих пор. Существует мощное научное течение - синергетика - которая пытается устранить указанное противоречие, опираясь на законы нелинейной неравновесной термодинамики. Согласно ее представлениям, в открытых системах, находящихся вдали от равновесия, энтропия может самопроизвольно уменьшаться, что и воспринимается нами как "упорядочение", "самоорганизация". Возникающие при этом структуры получили название "диссипативных" - в отличие от равновесных структур типа кристалла. Предполагается, что поскольку живые организмы суть открытые неравновесные системы, то и упорядоченность в них - диссипативного типа. Однако ни с логической, ни с фактической точек зрения этот вывод не представляется убедительным. Несомненно, диссипативные структуры в живых организмах существуют, но не играют там определяющей роли. Они занимают то же положение, что и другие физические и химические явления и механизмы - диффузия, осмос, теплопроводность, упругость, гравитация и т.д. То есть, там, где они способствуют достижению целей организма, им позволяют и даже помогают работать. Там же где они препятствуют достижению этих целей, они подавляются, блокируются, обходятся, компенсируются и т.п. По справедливому замечанию Л.А.Блюменфельда (1974), в живых системах слишком много жестких, практически не релаксирующих образований, устойчивость которых поддерживается высокими потенциальными барьерами, а отнюдь не термодинамическими силами. Мы бы добавили, что здесь еще больше параметров и структур, которые сохраняются за счет работы многочисленных систем регулирования и создаваемых ими управляющих сил, направленных на достижение целей системы, максимизацию ее целевой функции. Короче говоря, в организме гораздо больше от автоматически управляемой машины, чем от диссипативной структуры. Да и строится организм скорее, как машина, по определенному плану и программе, а не возникает в ходе спонтанных бифуркаций, в силу имманентной нелинейности, присущей строительному материалу, как мы видим в случае диссипативных структур. Таким образом, диссипативные структуры - это любопытные, но скорее всего локальные завихрения, возникающие на периферии двух мощных, но по-прежнему разнонаправленных потоков эволюции. Однако чтобы преодолеть "диссипативный" подход, нужно предложить взамен нечто столь же конструктивное, но более соответствующее сути биологической организации. Прежде чем перейти к этой сути, напомним еще одно интересное замечание Блюменфельда. "Так ли уж они упорядочены?" - спрашивает он по поводу живых систем. Ведь энтропия живого организма практически не отличается от энтропии куска горной породы того же веса. Отсюда, на наш взгляд, следует важный вывод, что интуитивно очевидная для нас высокая упорядоченность живых систем лежит где-то совсем в иной плоскости и вряд ли может быть уловлена и объяснена термодинамическими методами, по крайней мере, в их традиционном понимании. Какой же фактор, определяющий суть биологической организации, остается вне (или, по крайней мере, на периферии) поля зрения при традиционном термодинамическом подходе? Живой организм возникает в ходе адаптации, приспособления к условиям окружающей среды. Для биолога это - аксиома. Поэтому рассматривать процесс упорядочения и организации живой системы в отрыве от ее взаимодействия со средой - значит упускать из виду самую сущность этой организации. Упорядоченность живых организмов состоит в их целесообразности, приспособленности как целого к условиям среды, так и частей целого друг к другу. Равновесность или неравновесность возникающих при этом структур не являются определяющими. Имманентные свойства живой материи могут сформировать те или иные структуры, в том числе диссипативные, но только взаимодействие со средой будет определять, какие из этих структур полезны и должны быть сохранены при отборе. Именно на уровне макроскопических переменных, описывающих взаимодействие организма со средой, и происходит рост упорядоченности, определяющий "биологическое" направление эволюции. Для конструктивного описания этого процесса необходимо иметь количественную меру адаптированности и математический принцип, определяющий направленность протекания процесса. Желательно при этом, чтобы он охватывал, взаимоувязывал и объяснял обе тенденции эволюции. В ряде наших работ (Голицын, Петров, 1990,1991) было высказано предположение, что наиболее общей и адекватной мерой адаптации системы к окружению является средняя взаимная информация между условиями среды X и реакциями (или признаками)системы У . А основным принципом, определяющим эволюцию и поведение системы, является принцип максимума взаимной информации:
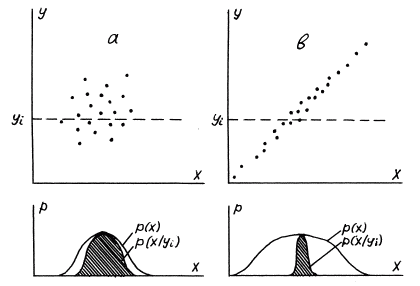
Здесь Принцип максимума информации означает, что по мере адаптации к среде признаки и реакции организма с все большей полнотой и точностью отражают условия среды. У хорошо приспособленного организма эта взаимная информация так велика, что биолог, наблюдая признаки организма, может довольно точно описать условия среды, в которой он обитает. И наоборот, зная условия среды, можно довольно правильно предсказать признаки организма, способного выжить в этой среде. Однако сказанное справедливо не только для живых организмов: любой предмет, сохранившийся в данной среде, неизбежно несет на себе более или менее отчетливый отпечаток этой среды, информацию о ней. Он вынужден в какой-то степени "приспосабливаться" к этой среде — в противном случае он просто погибает или превращается в нечто иное, более адекватное условиям среды. Так, кристалл соли не может выжить в воде, и если он сохранился, значит, среда была относительно сухой. Таким образом, адаптация - общее свойство материи, а принцип максимума информации, количественно описывающий это свойство, отражает общие тенденции эволюции материи. Для иллюстрации этого принципа рассмотрим некую среду с температурой Х ("термостат") и некоторый объект с начальной температурой У ("термометр"). В общем случае Х?У . Приведем теперь объект в контакт со средой. При этом начнется процесс релаксации, выравнивания температуры, в ходе которого энтропия системы "термостат + термометр", как известно, возрастает. Однако у этого процесса есть и другая сторона: его можно рассматривать как процесс адаптации объекта к среде, термометра к термостату. После завершения этого процесса, глядя на хорошо адаптированный термометр, мы можем судить о температуре термостата. Это значит, что возникла взаимная информация между двумя макроскопическими переменными - температурой термостата Х и термометра У . Конечно, мы давно знаем, что термометр может давать нам информацию о температуре среды и что для этого он должен быть сначала уравновешен с этой средой; однако редко задумываемся над тем, что здесь мы имеем примитивный прообраз более сложных и изощренных процессов биологической адаптации. (Кстати, И.П.Павлов по отношению к этим процессам употреблял именно термин "уравновешивание"). Дадим теперь более аккуратное описание этого процесса. Введем ансамбль из N одинаковых систем В ходе релаксации происходит "диагонализация" совместного распределения вероятностей Однако есть и другая тенденция. Система "термостат + термометр" находится в контакте с окружением и подвергается случайным воздействиям этого окружения. Подобно частице в броуновском движении, она может быть далеко уведена из своего начального состояния. Для ансамбля систем это означает диффузию, экспансию изображающих точек вдоль биссектрисы. При этом безусловное распределение вероятностей p(x) расплывается, безусловная энтропия H(X) возрастает и это, как видно из (1), также ведет к увеличению средней взаимной информации между X и У. Таким образом, в реальном процессе эволюции, как правило, наблюдаются обе тенденции - как рост энтропии (безусловной), так и организация, уменьшение энтропии (условной). Обе тенденции связаны с макроскопическими степенями свободы системы и, поскольку число этих степеней ничтожно по сравнению с числом микроскопических степеней свободы, то и вклад их в общую энтропию системы пренебрежимо мал. Однако, если в физических системах этим вкладом действительно пренебрегают, то в биологических системах он оказывается главным предметом нашего внимания. Литература
Рис. 1 Подписи к рисунку: Вверху: облако точек, изображающих состояния систем ансамбля. Внизу: распределения плотности вероятности: условной (сечение облака точек) и безусловной (проекция облака точек). Масштабы по вертикали условные: а. До релаксации: корреляция и взаимная информация отсутствуют. б. После релаксации: информация между Х и У возросла как за счет стягивания точек к биссектрисе, так и за счет растекания вдоль нее.
|